![]()
Videos from Maria Fischer, Stefanie Kaech, Darko Knutti & Andrew Matus, 1998, "Rapid Actin-Based Plasticity in Dendritic Spines". Neuron 20, p. 847-854.(Downloaded from http://www.neuron.org/cgi/content/full/20/5/847/DC1) |
|
|
|
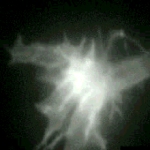
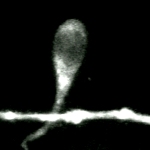
Growth cone on a 48-hr-old hippocampal neuron transfected with GFP-actin. In addition to the concentration of actin in the "palm" of the growth cone, local concentrations are associated with spots and lateral filopodia from the shaft of the neurite. Time marker shows hours, minutes, and seconds. |
|
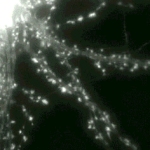
A large field from a GFP-actin expressing hippocampal neuron is shown at low magnification (40X objective lens). As well as showing the widespread nature of spine motility, this sequence also indicates the extent to which actin is concentrated in spines compared to the shaft domain of dendrites. Time marker shows hours, minutes, and seconds. |
|
|
|
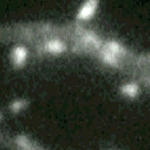
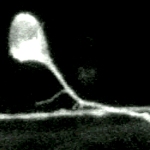
Dynamics in motile spines recorded at higher magnification (100X objective lens) from a GFP-actin transfected cell in another culture. Over these short recording times the actin-driven changes are limited to spine shape and involve the growth and shrinkage of miniature protrusions. Time marker shows hours, minutes, and seconds. |
![]()
Videos from Noam E. Ziv & Stephen J. Smith, 1996, "Evidence for a Role of Dendritic Filopodia in Synaptogenesis and Spine Formation", Neuron 17, p. 91-902.(Downloaded from http://www.neuron.org/cgi/content/full/17/1/91/DC1) |
|
|
|
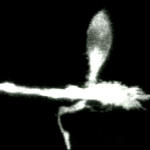
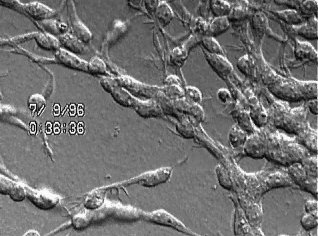
The video begins with a low magnification Nomarski image of a pyramidal neuron raised in culture for 11 days. The next segment of the video shows the neuron has been labeled with the fluorescent lipophilic dye FAST DiO. The final video segment contains a time-lapse sequence composed of 20 frames taken at 3 minute intervals showing the rapid morphological changes of the labeled dendrite and its filopodia that occurred over a period of ~1 hour. This sequence is shown both at fast and slow playback rates. |
![]()
Videos from James D. Jontes, JoAnn Buchanan & Stephen J. Smith , 2000, "Growth cone and dendrite dynamics in zebrafish embryos: early events in synaptogenesis imaged in vivo", Nature Neuroscience 3-2, p. 231-237.(Downloaded from http://www.nature.com/neuro/web_specials/) |
|
|
|
Simultaneous two-photon imaging of the Mauthner growth cone and primary motor neurons. The video sequence shows maximum-intensity projections from a representative time-lapse sequence during which an M-cell growth cone migrated past a CaP motor neuron. The growth cone was highly active and transiently interacted with the CaP cell, one of its synaptic partners. The growth cone moved past the CaP cell without collapsing, stalling or significantly altering its morphology. |
|
Formation of an axonal varicosity. This two-photon time-lapse sequence shows the formation of an axonal varicosity in close association with a CaP motor neuron. The images are maximum-intensity projections of image stacks displayed at five-minute intervals. Initially, the axon swelled, forming a large, elongated varicosity close to the CaP cell. Over time, the varicosity shrank and became more spherical. Throughout the time-lapse, there was extensive filopodia activity on the CaP cell near the developing varicosity. |
|
|
|
Interaction of dendritic filopodia with the Mauthner axon. This two-photon time-lapse sequence illustrates the interaction of a Mauthner growth cone with a RoP motor neuron as these two cells first came into contact. At the start of this time-lapse sequence, the Mauthner growth cone first reached the RoP ventral dendrite. Each image is a maximum-intensity projection of 14 sections collected at 1-mm steps. Numerous dendritic filopodia extended and retracted from the RoP ventral dendrite, many of which interacted transiently with the Mauthner axon. During this experiment, no stable cell-cell contacts were observed. Because a synapse will ultimately form between the Mauthner axon and the RoP ventral dendrite, it is possible that a filopodium may be responsible for initiating synaptogenic contact. |
Videos from Hynek Wichterle, Jose Manuel García-Verdugo & Arturo Alvarez-Buylla (1997), "Direct Evidence for Homotypic, Glia-Independent Neuronal Migration.", Neuron 18, 779–791.(Downloaded from http://www.neuron.org) |
|
| Neuronal precursors born in the subventricular zone (SVZ) of the neonatal and adult rodent brain migrate 3–8 mm from the walls of the lateral ventricle into the olfactory bulb. This tangentially oriented migration occurs without the guidance of radial glia or axonal processes. The cells move closely associated, forming elongated aggregates called chains, which are ensheathed by astrocytes. We have developed a culture system in which postnatal mouse SVZ neuronal precursors assemble into chains with ultrastructural and immunocytochemical characteristics equivalent to those in vivo but without the astrocytic sheath. Time-lapse videomicrography revealed that individual cells migrate along the chains very rapidly (~122 µm/hr) in both directions. Periods of cell body translocation were interspersed with stationary periods. This saltatory behavior was similar to radial glia–guided migration but ~4 times faster. Neuronal precursors isolated from embryonic cortical ventricular zone or cerebellar external granule layer did not form chains under these conditions, suggesting that chain migration is characteristic of SVZ precursors. This study directly demonstrates that SVZ neuronal precursors migrate along each other without the assistance of astrocytes or other cell types. (Additional data are presented in www.cell.com). | |
Video from Nadarajah, P Alifragis, R O L Wong & J G Parnavelas (2002) "Ventricle-directed migration in the developing cerebral cortex B" Nature Neuroscience 5, 218 - 224.(Downloaded from http://www.nature.com/neuro/web_specials/) |
|
|
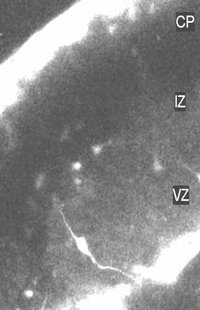
It is believed that postmitotic neurons migrate away
from their sites of origin in the germinal zones to populate distant targets.
Contrary to this notion, we found, using time-lapse imaging of brain slices,
populations of neurons positioned at various levels of the developing
neocortex that migrate towards the cortical ventricular zone. After a
pause in this proliferative zone, they migrate radially in the direction
of the pial surface to take up positions in the cortical plate. Immunohistochemical
analysis together with tracer labeling in brain slices showed that cells
showing ventricle-directed migration in the developing cortex are GABAergic
interneurons originating in the ganglionic eminence in the ventral telencephalon.
We speculate that combinations of chemoattractant and chemorepellent molecules
are involved in this ventricle-directed migration and that interneurons
may seek the cortical ventricular zone to receive layer information.
|
|
![]()
Video from Borgdorff AJ & Choquet D. (2002) "Regulation of AMPA receptor lateral movements." Nature 417(6889): 649-653.(Downloaded from http://www.nature.com/nature/journal/v417/n6889/suppinfo/nature00780.html) |
|
| video recording at 5x real time of the movement of two GluR2 bound 0.5 µm latex beads at the surface of the neurite of a 2 DIV hippocampal neuron | |
| video recording at 5x real time of the movement of a GluR2 bound 1 µm latex beads at the surface of the neurite of a 3 DIV hippocampal neuron in the presence of a caged calcium ionophore. At the red spot, a train of U.V. flashes was applied around the bead to trigger a local increase in intracellular calcium level. | |
| video recording at 5x real time of the movement of two GluR2 bound 0.5 µm latex beads at the surface of the neurite of a 14 DIV hippocampal neuron. Note that both beads stop and resume diffusive movement at the same time. Movie form the experiment shown in Fig. 2. | |
Videos from dowloaded from http://laxmi.nuc.ucla.edu:8888/Libraries/Animations |
|
|
|
|
|
|
|
|
|
|
| The brain | Main menu | Studying the brain |